

Biophysics of Cell Movements
By: Arpita Upadhyaya
Cells are the basic units of life and all our genetic material is housed inside them. Cells are bursting with biochemical activity as proteins assemble and go about their life-giving work. They also find time to divide and multiply as well as move from one place to another. The motivation to study movements of and within living cells may be as simple as that which led Leeuwenhoek to peer down the first microscope several hundred years ago and exclaim in wonder at the various different types of moving things he saw. In spite of their genetic and biochemical complexity we can think of cells as physical systems that are bound by physical laws. Nature has engineered many molecular and cellular machines that induce motion at different scales – from the level of a single molecule to that of multi-cellular tissues. One challenge for physicists is to understand how biology exploits physical principles to cause movement.
Motion in microscopic living organisms assumes diverse forms – such as swimming of bacteria using a rotary motor, the spring-like contraction of Vorticella, the rapid beating of tiny hairs (cilia) in Paramecium to name a few. The common theme in all cell movement is that chemical energy is converted to mechanical work and motion. There are a number of ways in which this can be achieved. Just as in our everyday world, propellers, motors, springs and pistons store or release energy to power motion, their analogs inside cells can perform similar functions in biological systems. However, unlike the familiar springs and motors which are macroscopic, their cellular counterparts are typically nanometers (10 -9m) to microns (10-6 m) in size. Such objects are constantly buffeted by collisions with water molecules due to thermal fluctuations. Another point to consider is that the typical velocity scales of these objects is at best 10-6 m/sec, implying that inertia is negligible and viscous effects dominate. At physiological temperatures, thermal motion is significant and therefore the viscous and fluctuating cellular environment constrains the modes of motion generated and the kinds of forces that are required.
 |
|
| Figure 1: The top panel shows a keratocyte cell as it crawls on a substrate. The cell is about 10 microns in diameter. The bottom panel shows an electron microscope image of a small portion of the cell “leading edge” |
Much research is aimed at understanding one of the most common kinds of movement - the crawling motion of cells which is essential for organism development and survival. For example, cells of our immune system move towards the site of infection, and groups of cells change shape and coordinate their movement to repair wounds or construct organs during development. Improper regulation of the mechanical behavior of cells has been linked to a number of diseases, including asthma, cardiac arrhythmia and cancer metastasis. If we look at individual crawling cells through a microscope, we are struck by the dynamic changes in cell shape and location. These are mediated by the cytoskeleton, a complex array of proteins that act as the scaffold of cells. The most abundant protein of the cytoskeleton is actin which tends to form long filaments that are organized near the cell membrane and provides much of the force required for cell movement. While the mechanical properties of the cytoskeleton gives cells their rigidity, its assembly and disassembly plays a crucial role in helping cells to move. Cytoskeletal filaments such as actin are polymers that can spontaneously elongate by addition of monomers in a process called polymerization. How can the growth of protein filaments by addition of monomers cause motion?
Terence Hill and subsequently, George Oster and Charlie Peskin realized that this process of polymerization can be used to generate mechanical forces that can propel objects. Suppose we consider a filament growing at one end by addition of subunits or monomers. If it encounters a load in front of it, it may not be able to directly ‘push’ it as there is not enough room for the addition of the next subunit. However, because of thermal motion, the load and the filament can both fluctuate. If the load is fluctuating, (the size of the fluctuations being typically larger than each monomer), the diffusing monomer finds some space to attach at the end of the filament. Thus, the filament has grown longer and the load can no longer move back to its original position. Thus, the addition of a monomer has effectively “ratcheted” the load forward in one direction – in other words it acts as a “Brownian ratchet”. However, we have not gained something for nothing, since energy from ATP hydrolysis must be expended in order to keep a supply of monomers available. We see that polymerization can act as a “motor” to propel micron sized objects against load forces of several piconewtons (10-12 N).
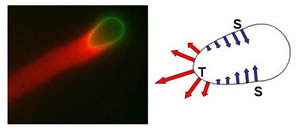 |
|
Figure 2: Bio-mimetic system of actin (stained with a red fluorescent dye) and phospho-lipid bilayer vesicles (green). Actin filaments form a “comet” tail as they polymerizing against the vesicle and propel it forward. The forces generated by actin polymerization (as shown by the arrows) can be deduced by quantifying the deformations of vesicles with known mechanical properties. The vesicle shown is about 4 micron in diameter |
Long single filaments (micron sized) are quite flexible and can buckle easily under the effect of piconewton forces. But cells do not have single actin filaments in isolation. From electron microscopy images we can see that the cell has a dense meshwork of filaments (Figure 1). There are many proteins in the cell that bind actin to cross-link individual filaments into large networks. The unsupported ends of these filaments are only a few hundred nanometers in length, and therefore can sustain much larger forces. The structural and mechanical properties of these gel-like networks depend on the type and concentration of the actin-binding proteins. Further, actin networks, unlike many other cell components, alter their growth in response to forces, not just chemical signals. Such cytoskeletal material provides quite a number of interesting problems in “soft” condensed matter physics. In contrast to traditional rubber-like networks, these polymers are semi-flexible and the energy required to bend the filament on micron length scales is comparable to thermal energy. The competition between enthalpic and entropic effects in the dynamics and deformation of semi-flexible networks lead to extremely rich and varied mechanical response of both entangled solutions and networks cross-linked by actin-binding proteins.
How does this labile, self-organizing system of filaments, membranes and associated proteins conspire to drive directed motion of cells in response to a diverse array of environmental cues and internal signals? How do cells generate forces, respond to forces and sense their physical environment? We develop tools to push, pull, squeeze, tug and observe the dynamic structure and biophysical behavior of cytoskeletal filament assemblies at sub-micron length scales to study how cells generate and transmit mechanical forces. These small structures require tools such as optical and magnetic traps to manipulate them and powerful microscopes to observe them. Another way to understand the workings of the actin cytoskeleton is to build artificial “cells”, or bio-mimetic systems, from the parts list of membranes, action and associated proteins as shown Figure 2. This way we can examine the function of different parts and how it fits into the full “polymerization motor”. Ultimately, we would like to be able to relate the forces generated by actin networks under artificial conditions to those inside living cells. To this end, we use high resolution fluorescence microscopy to observe cytoskeletal protein dynamics in living cells and, simultaneously, measure their biophysical properties at micron length scales. Future work in our biophysics lab at the University of Maryland will involve using various imaging, genetic and micro-manipulation techniques to uncover the physical basis of biological motion.
--------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------
Dr. Upadhyaya is an Assistant Professor at the University of Maryland Department of Physics. She is a member of the Biophysics Group. For any questions or comments, please contact her at arpitau@umd.edu.